
simPEATC, es un proyecto de simulador de PEATC, para estudiantes de electrofisiología auditiva, en el cual se intenta recrear el procedimineto completo
- [:speak_no_evil:]GUI
- [:speak_no_evil:]Insertar Matplotlib seteado
- [:construction_worker:]i18n
- [:rat:]Configuración de tonos
- [:speak_no_evil:]Desarrollo del Hadware
🆗:OK, 🙊:50/50, 👷:en construcción, 🐀: nivel rata(no funciona, ¡aún!)
02-10-2019:
- Se agregan stickers para preamplificador y portaelectrodos en formato SVG de Inkscaper
- Se modifica orden de las carpetas y archivos para una mejor interpretación
- Se comienzan a crear los archivos FZZ de fritzing para mostrar de forma sencilla los cirtuitos internos del preamplificador y portaelectrodos
 |
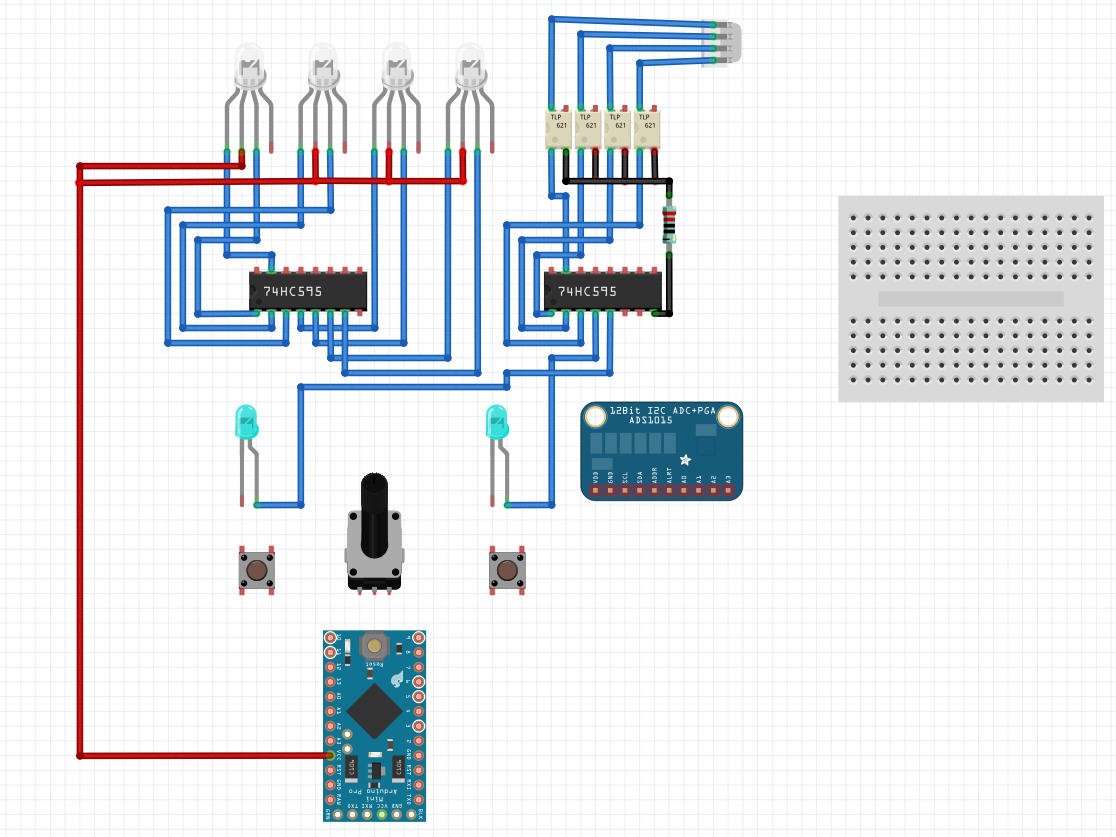 |
| Stickers | Fritzing - Preamplificador |
01-10-2019:
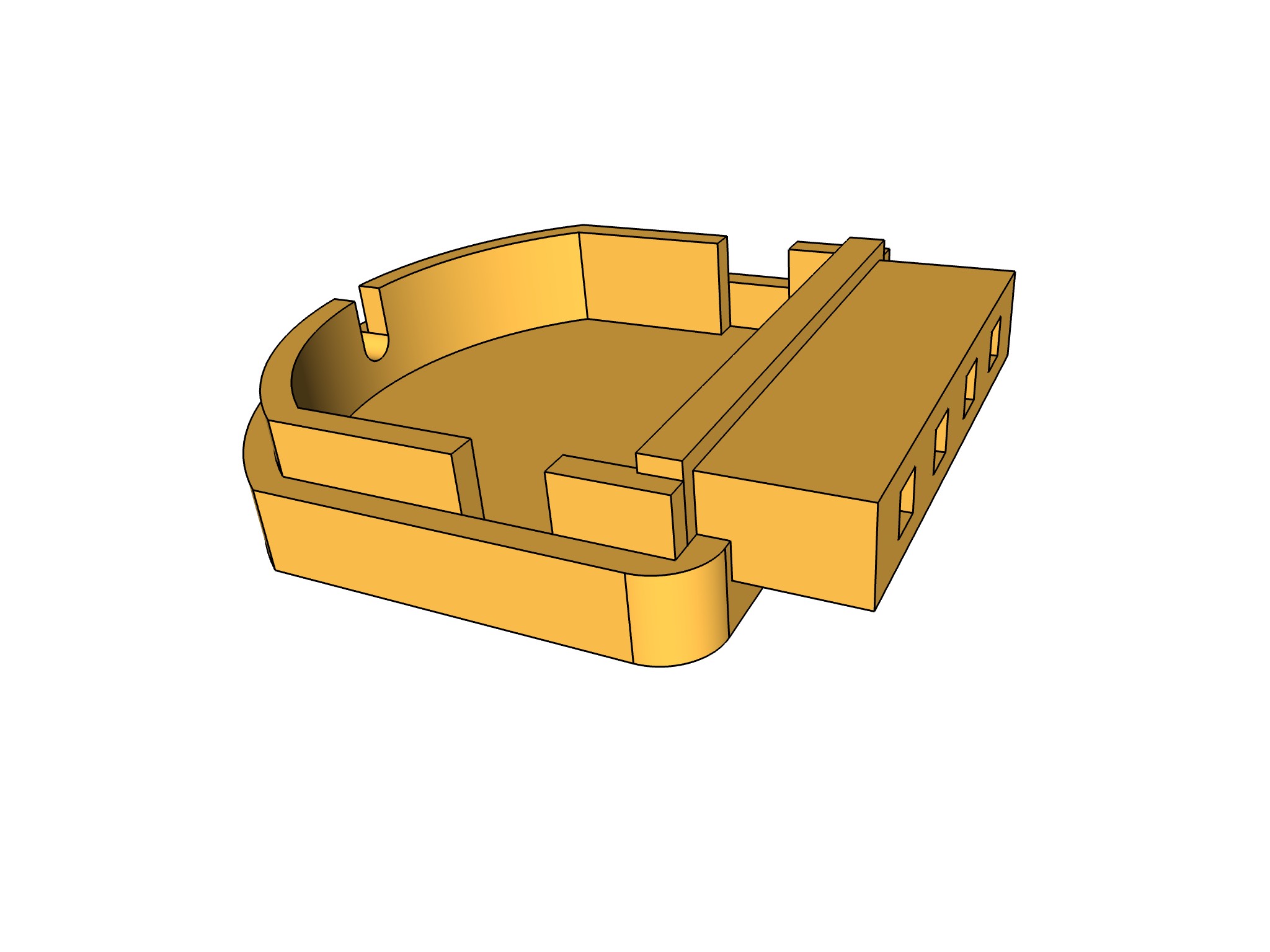
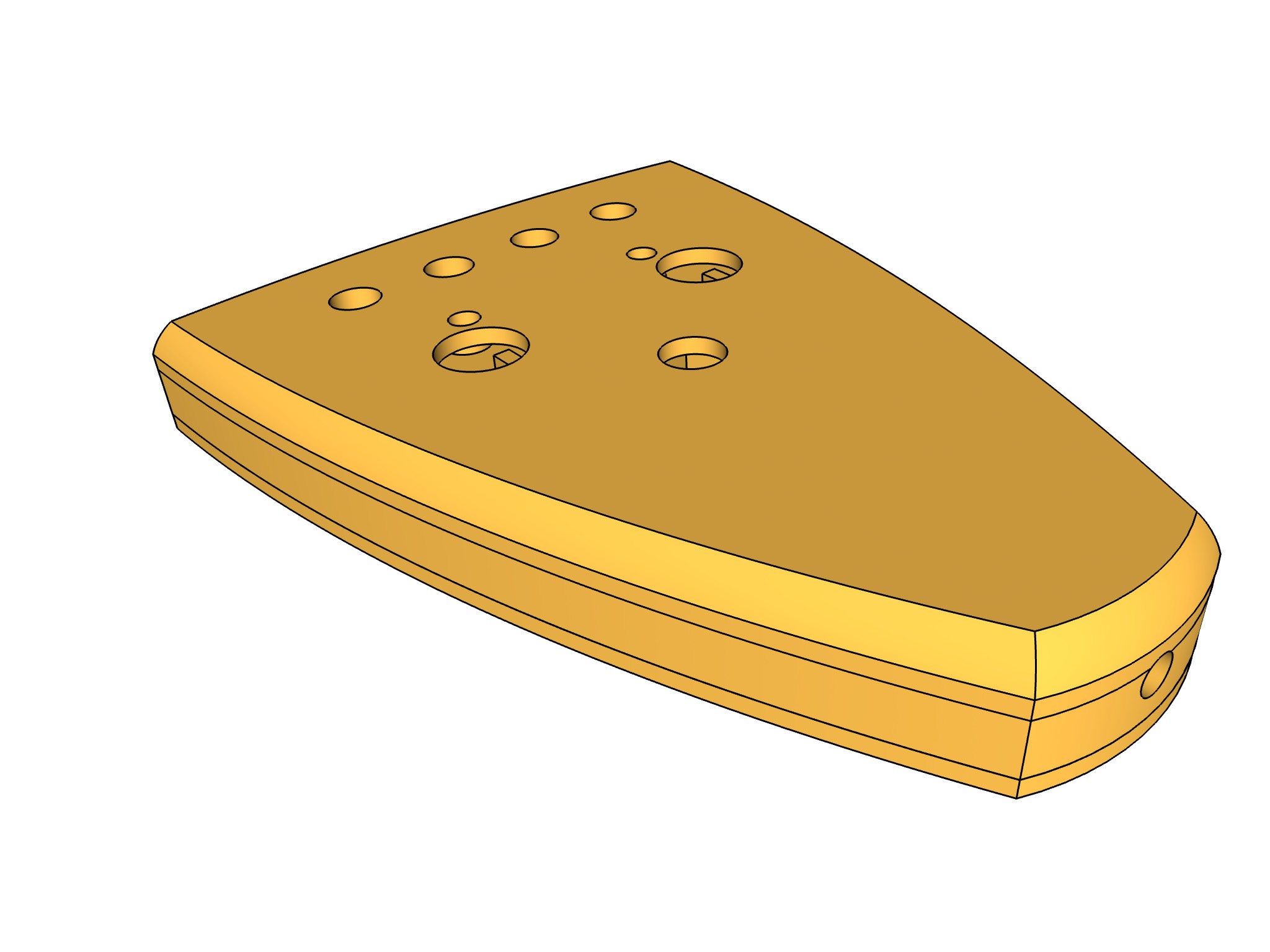
- Se agregan formatos STL, para impresión en 3d del preamplificador y portaelectros (carpeta STL): Se construyo el porta electrodos para poder fabricar electrodos con pin dupont de 1 pin
 |
 |
 |
| porta electrodos | preamplificador | impresión 3d |
08-09-2019:
- Se consigue generar animación de la formación de los potenciales evocados dentro de la gui.
bugs: el sistema queda en un bucle de matplotlib impidiendo realizar acciones mientras se genera el potencial

02-09-2019:
- Se realiza gui completa de la pantalla principal
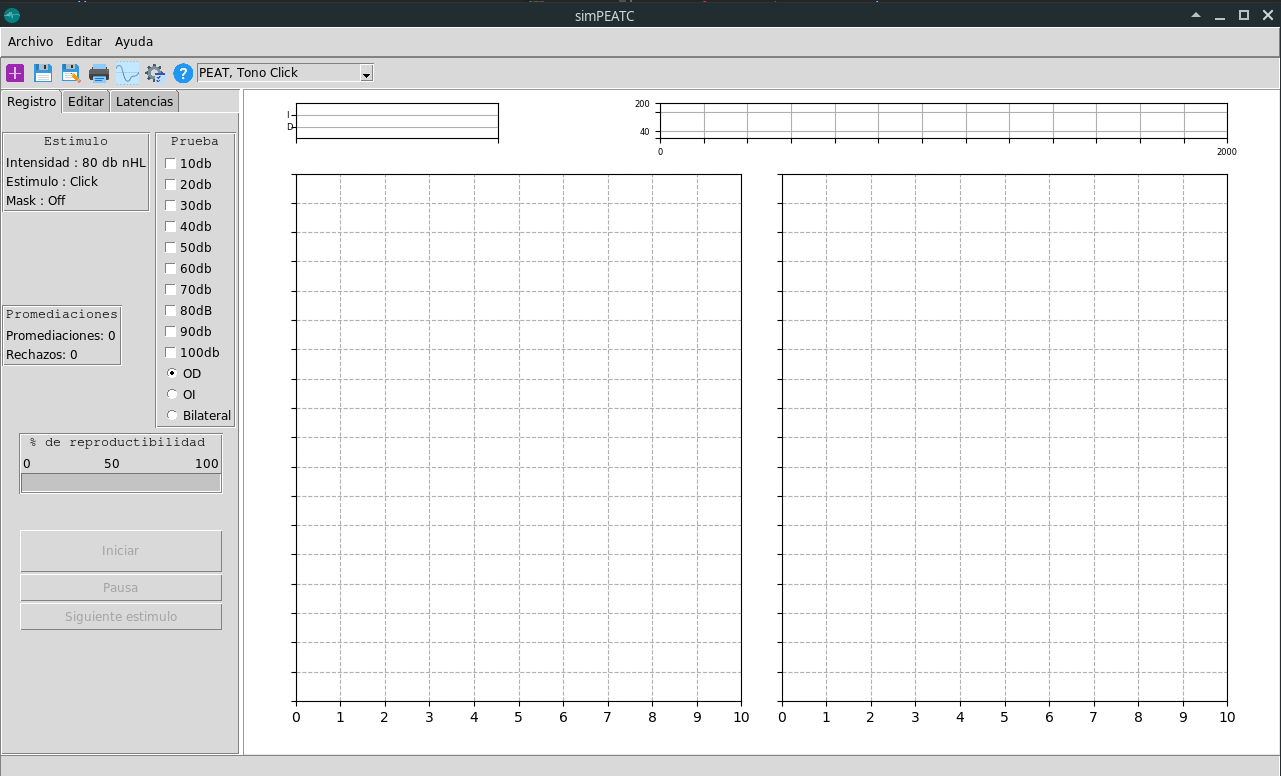
Para correr este script de Python es necesario tener instalado:
Python 3.x
Matplotplib
Tkinter
PIL
Numpy
csv
| Paquete | Código | |
|---|---|---|
| Linux | ||
| windows |
Este proyecto se ha desarrollado con las siguientes herramenientas
 |
David Ávila Quezada
|
 |
María Paz Latorre Gonzalez
|
 |
Ignacia Inarejo Inarejo
|
Este proyecto está bajo la Licencia MIT - mira el archivo LICENSE.md para detalles
- Escuela de Tecnología Médica Universidad Austral de Chile - Sede Puerto Montt - TM-PM