Engineering Bacterial Colonies for Advanced Gaming and Bidirectional Communication with Human Brain Organoids



We want to create bacteria colonies that will be able to play Pong, Go, DOOM, and to decode/encode data to/from human brain organoids
In the rapidly evolving sphere of bioelectrical cell communication, advances in bioengineering have paved the way for new insights into microbial behavior, including bacterial biofilms. A bold research proposal presented in this article merges principles of bioelectricity and artificial intelligence (AI), intending to construct a 'bacteria brain.'
This innovative research aims to harness bacterial bioelectric signals to emulate AI's data processing within a bacterial colony, transforming the colony's environmental perceptions into problem-solving abilities. The primary goal is to create a bacteria colony capable of interacting with games like Ping-pong and Go, by converting their bioelectric signals into game commands, thus setting up a dynamic feedback loop.
This groundbreaking research postulates that such a loop could foster the development of information processing pathways, akin to those in neuronal systems. The integration of these principles with mathematical models could unveil a universal algorithm of biological systems, crucial for understanding complex information processing.
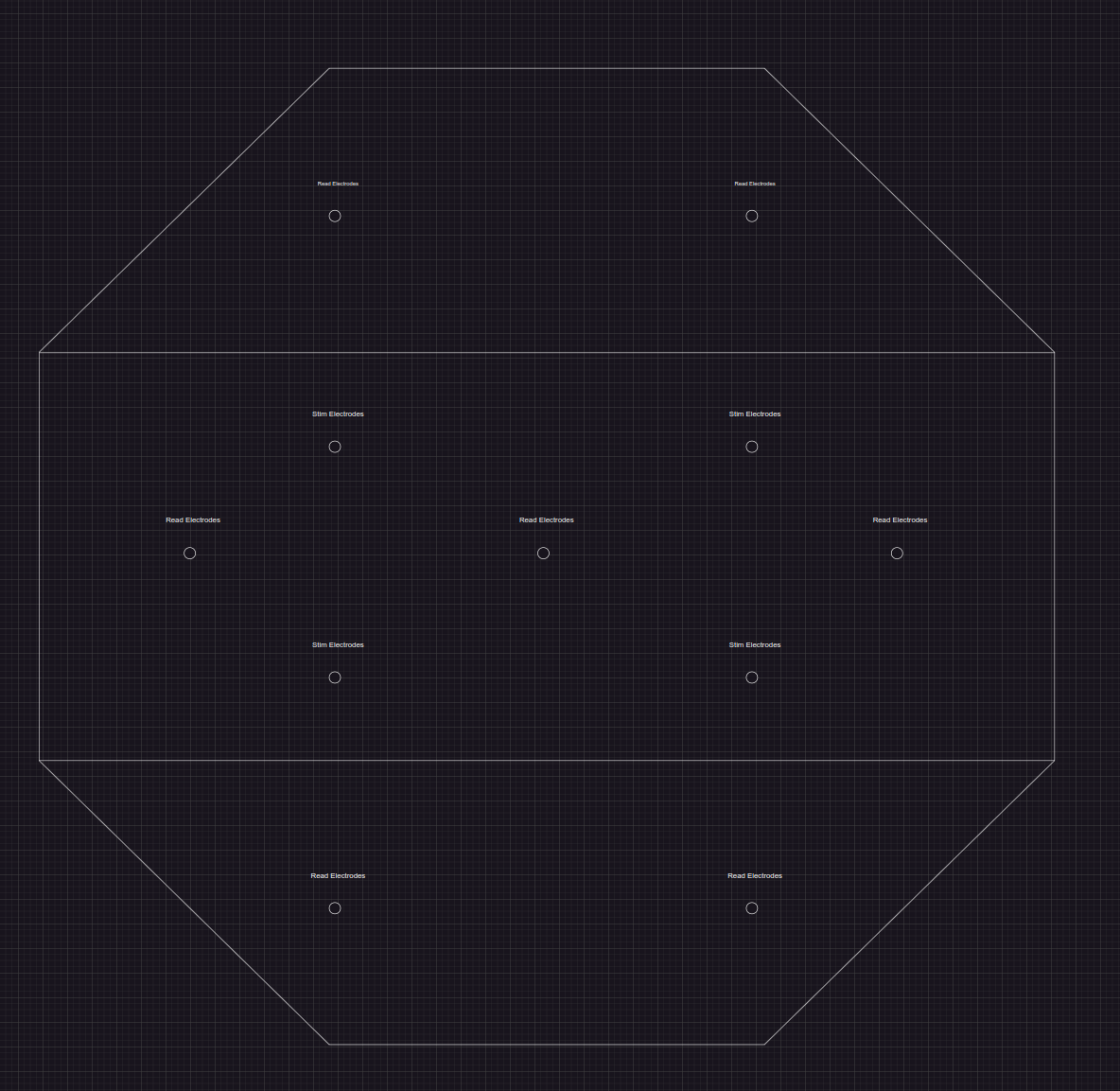
The meticulously designed experimental setup involves a controlled environment that accurately tracks and records the bioelectric signals from a bacterial colony, advancing our understanding of cell-to-cell communication mechanisms.
A unique reward system utilizing microfluid nutrients and electrical stimulation will be implemented in the experimental design, following the Free Energy Principle that suggests adaptive organisms resist disorder. These nutrients and electrical stimulations will reinforce specific bioelectric signal patterns, fostering beneficial signal generation.
In this setup, the 'bacteriabrain' is expected to reduce signal chaos, aiding in network organization. By inducing predictable, random, and sensory stimuli, the bacterial network is trained to adapt and learn, potentially leading to the development of advanced bioelectric signal processing capabilities.
In conclusion, this pioneering research has the potential to significantly contribute to the development of a neural link composed entirely of bacteria, marking a departure from traditional designs using conventional circuitry or neurons. By harnessing the inherent bioelectric capabilities of bacteria, we may be able to construct a more adaptive, resilient, and energy-efficient neural link. The bio-based system may offer superior biocompatibility, reducing the risk of immune responses often seen with traditional synthetic interfaces. Moreover, the bacterial system's potential to self-repair and replicate could vastly outperform current neural link technologies. The findings from this groundbreaking research could thus not only reshape our understanding of biological information processing but also propel us into a new era of bio-engineered neural interfaces.
We're more than a project; we're a movement. Let's make history together. Get in touch!
This project is licensed under the Creative Commons Attribution-ShareAlike 4.0 International License.
- Bio Silicon Synergetic Intelligence System: Combining Rat Brains and AI to create bio silicon intelligence
- Human Cortical Organoid Signal Analysis: Signal analysis and prediction of brain signals, adaptable for various signal types. PyPI libraries available from our experiments.
- EEG Prediction with Chaos Theory: Leveraging chaos theory and a CNN Kuramoto transformer RNN for signal prediction. Includes PyPI library implementations.
For collaborations, press inquiries, or questions:
- Email: soul.syrup@yandex.com or soul.syrupp@gmail.com
- Discord: soul_syrup
We invite you to test our PyPI library for human brain cortical organoid/spheroid, EEG, ECoG, and other signal analyses:
- Neural Signal Analysis Library from Synthetic Intelligence Labs.
Bacteria Neural Network by Synthetic Intelligence Labs is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Based on a work at Synthetic Intelligence Labs.
Permissions beyond the scope of this license may be available at Synthetic Intelligence Labs.
