{kind=link}
{kind=link}
1- Clone the repository
> git clone git@github.com:ealtamir/pf_experimentos.git2- Create command line tool on XCode, setting the language on C++
3- Delete the main.cpp on the created project
3- Drag and drop the folder src to the project and select the "Create Groups" options
4- Drag and drop the folder include to the project and select the "Create Groups" options
5- Move the main.cpp on src folder to the where main.cpp was on the step 3
6- Compile libraries from bullet3 (https://github.com/bulletphysics/bullet3)
Compile libraries on OSX:
> git clone git@github.com:bulletphysics/bullet3.git
> cd bullet3
> cmake . -G "Unix Makefiles" -DINSTALL_LIBS=ON -DBUILD_SHARED_LIBS=ON \
-DFRAMEWORK=ON -DCMAKE_OSX_ARCHITECTURES='i386;x86_64' \
-DCMAKE_BUILD_TYPE=RelWithDebInfo -DCMAKE_INSTALL_PREFIX=/Library/Frameworks \
-DCMAKE_INSTALL_NAME_DIR=/Library/Frameworks -DBUILD_DEMOS:BOOL=OFF
> make -j4
> sudo make install7- Drag and drop the following libraries installed on '/Library/Frameworks/' to the root of the project: -BulletDynamics.framework -BulletCollision.framework -LinearMath.framework
7.bis- Installing galib:
-Get the path to the include folder of the galib, '$REPOSITORY_PATH/libraries/galib/247/include'
-Go to xcode, select the project target and add that path to the 'header search path'
-Also add the libga.a dragging it to the root target of the project on xcode ( the libga.a is inside '$REPOSITORY_PATH/libraries/galib/247/lib/libga.a' ).
In case the static library don't work do the following:
brew install galib
and find the path where it was installed(usually /usr/local/Cellar/galib). The folder tree will be like the one described above to find the static lirary
8- Drag and drop the static libs on the folder 'libraries' to the root of the project
9- Select the root of the project, go to build phases, open 'Link Binary With Libraries' and add: -GLUT.framework -OpenGL.framework
10- Select the root of the project, go to build phases, open 'Compile Sources' and add all the .cpp files
After all you should get something like this:
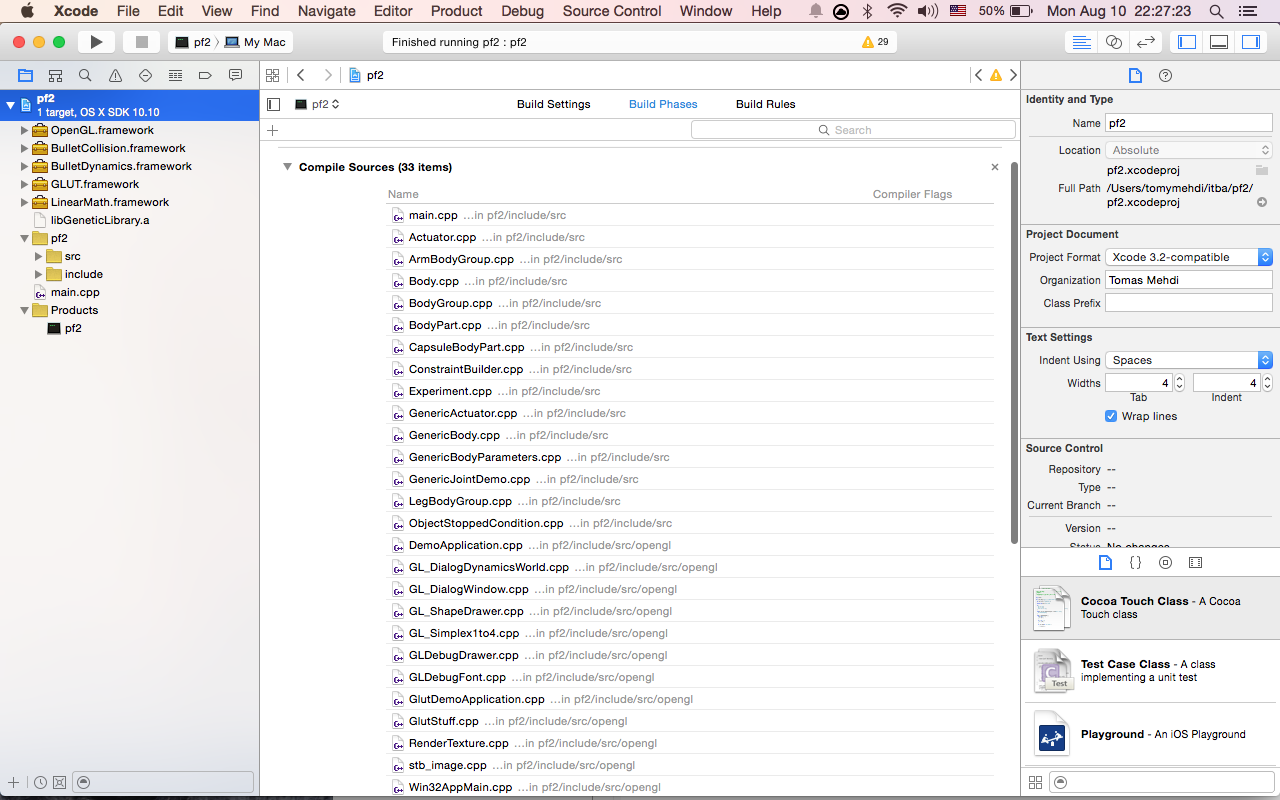
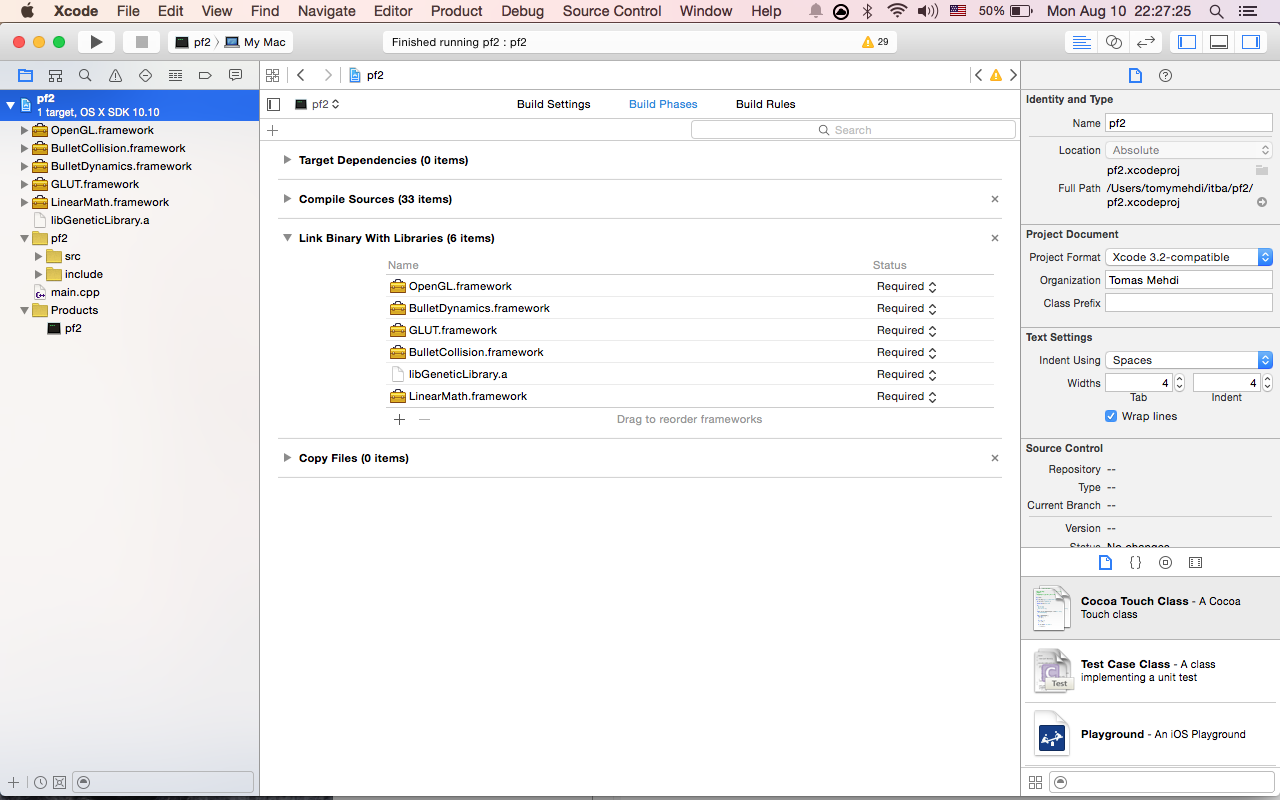
11- Change the XCode's build location. This is needed because we use relative paths to read files from the project directory. In order to do this, you have to go to XCode's options (shortcut Command-,), then Locations > advanced button, choose custom and enter something like: "/Users/Username/PF/proyectoDir/bin" for intermediates enter "/Users/Username/PF/proyectoDir/bin/Intermediates". Donde proyectoDir es el directorio del repositorio de github.
- Followed all the steps, tried to compile the project and failed because errors in one specific file. Try to remove all files from the compile sources list and add them again. Maybe you missed an important file the first time. Remember to clean de project before retrying compilation.
This project is maintained by Tomás Mehdi, Enzo Altamiranda and Teresa Fontanella De Santis.