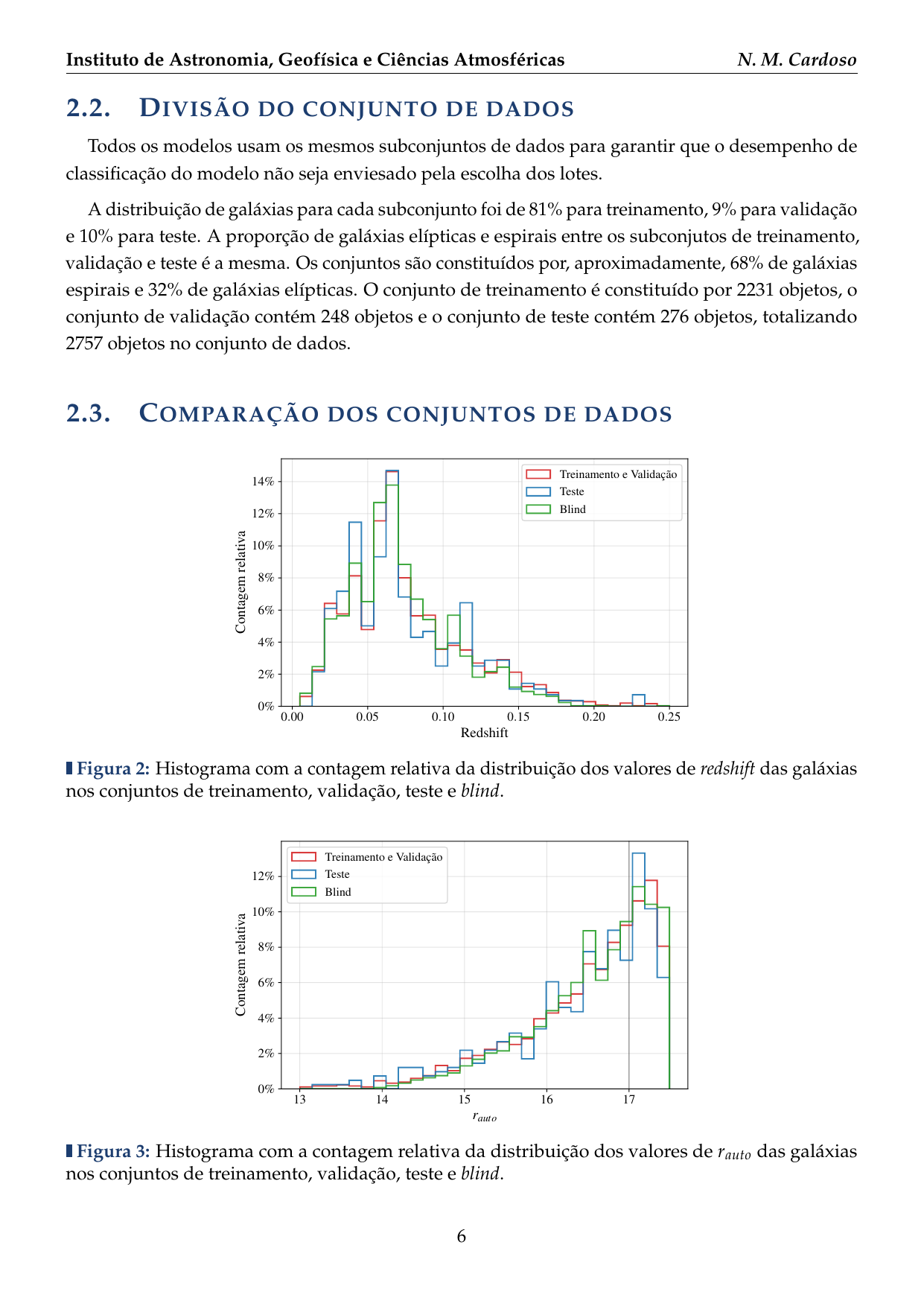
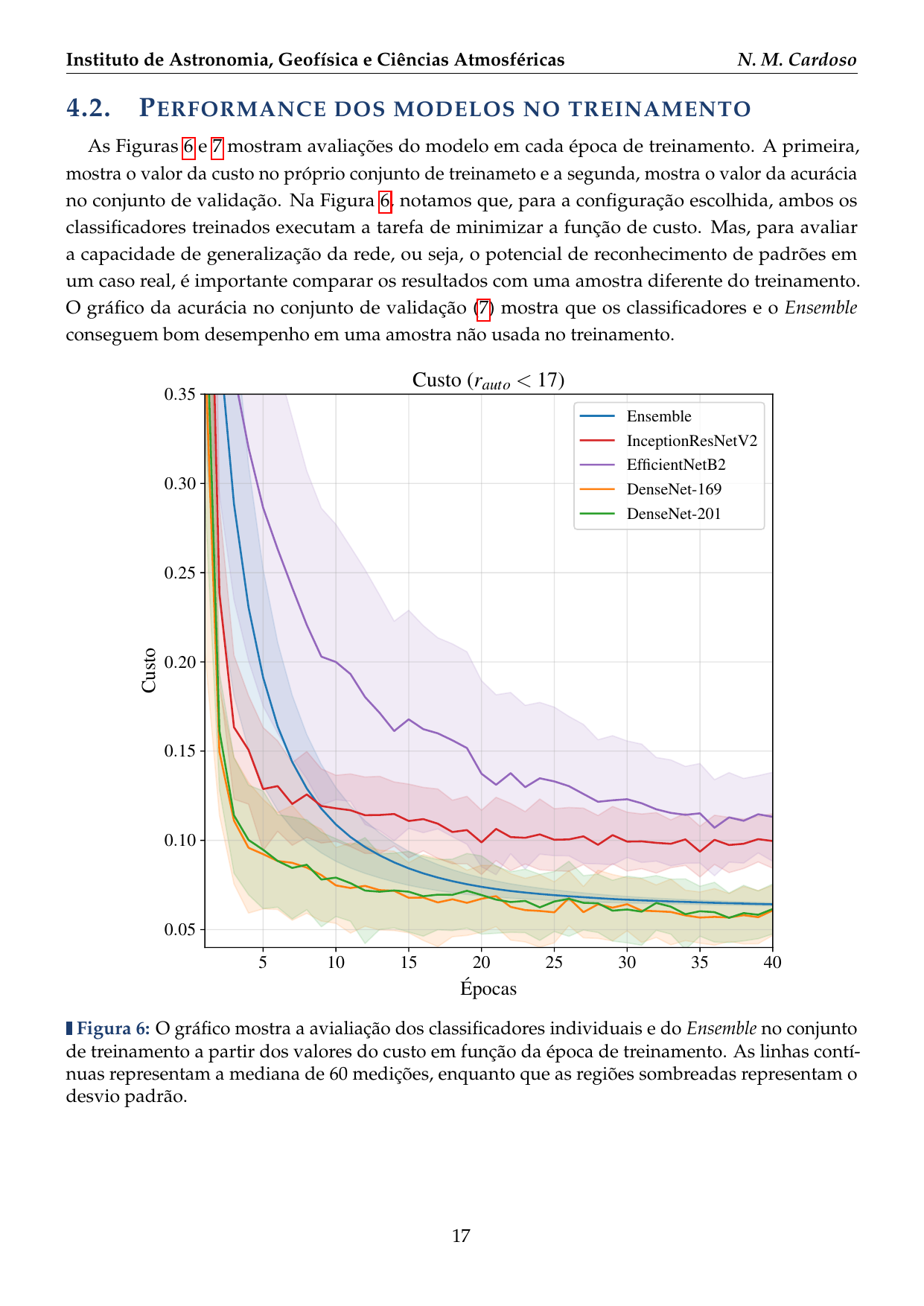
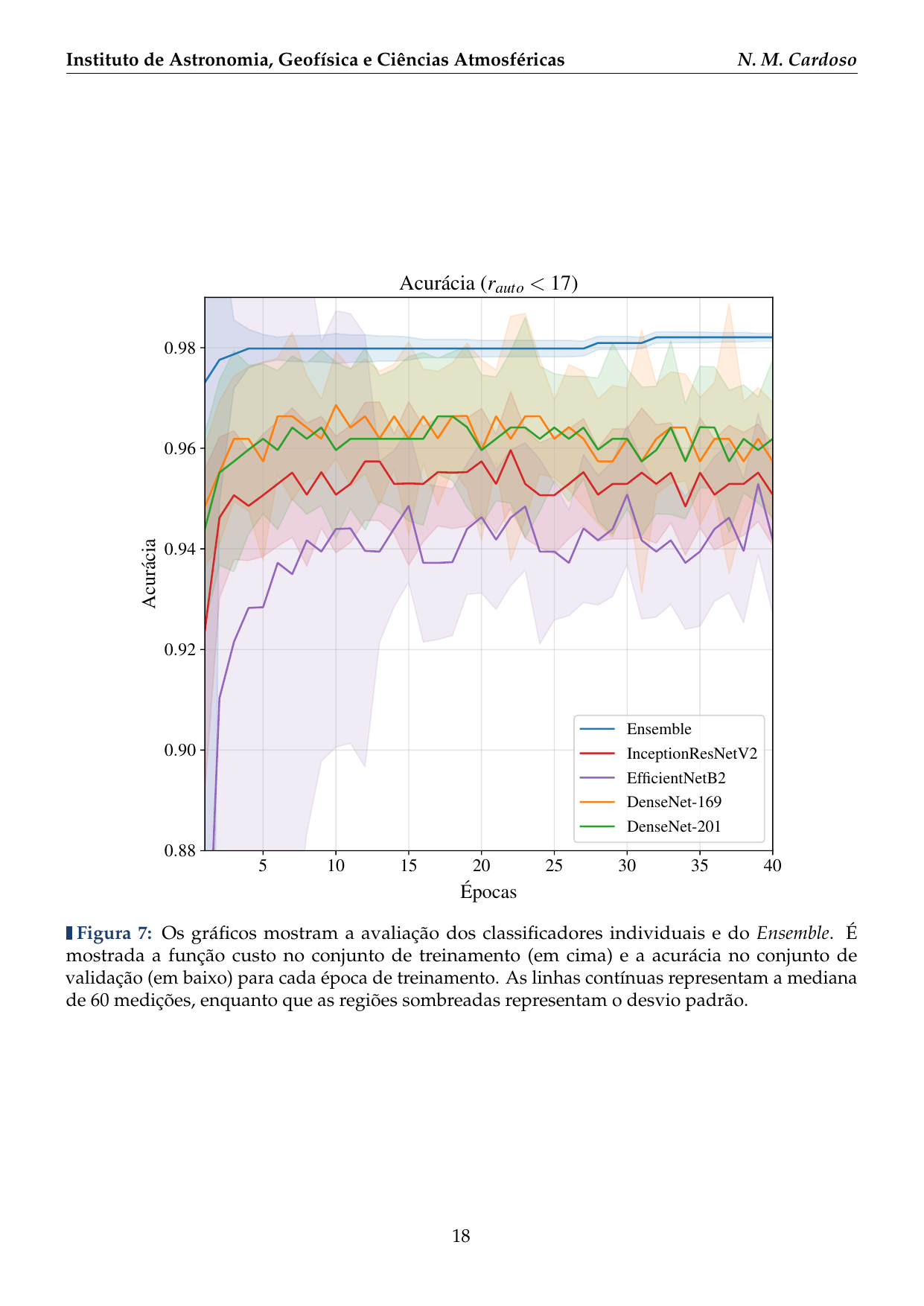
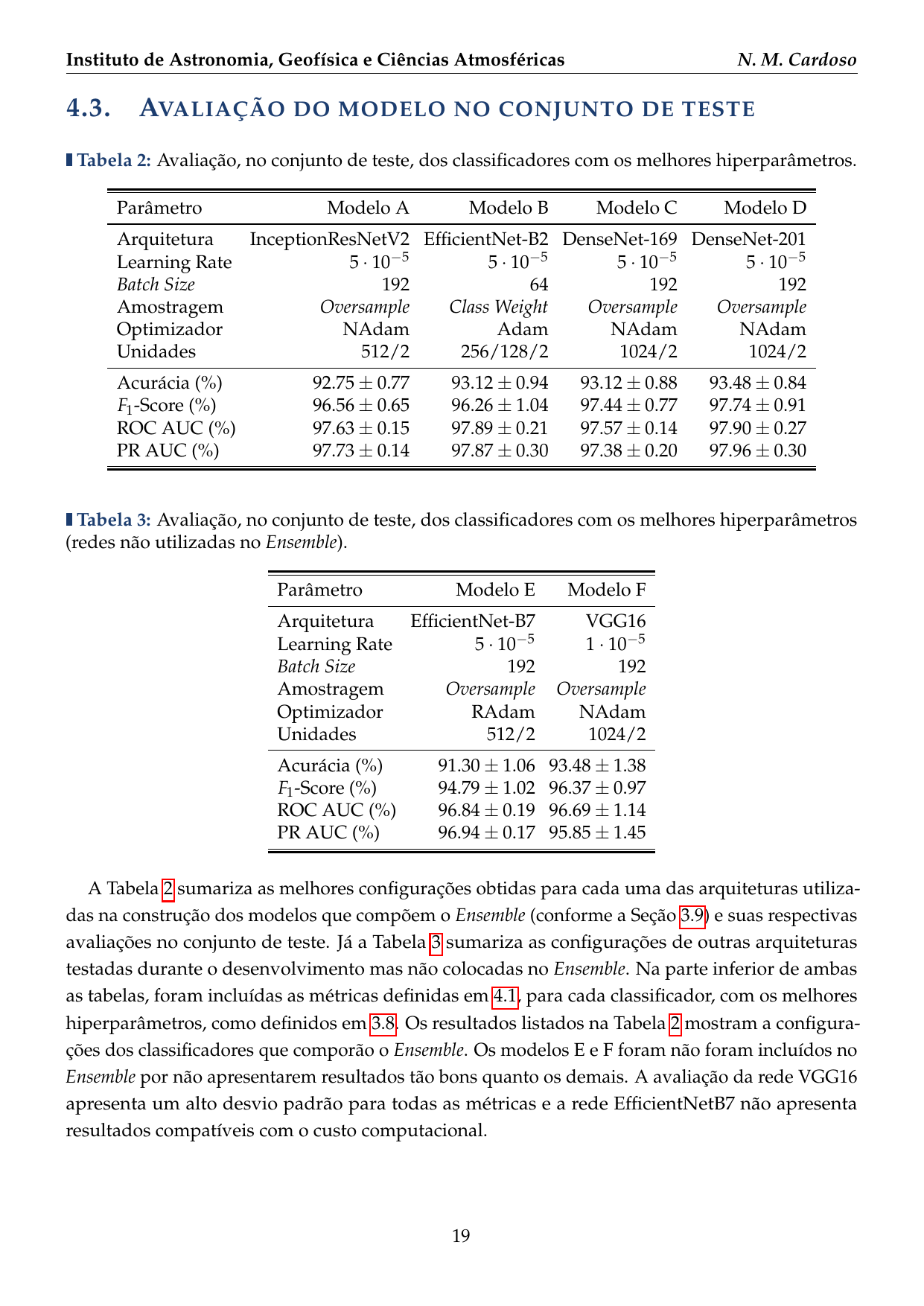
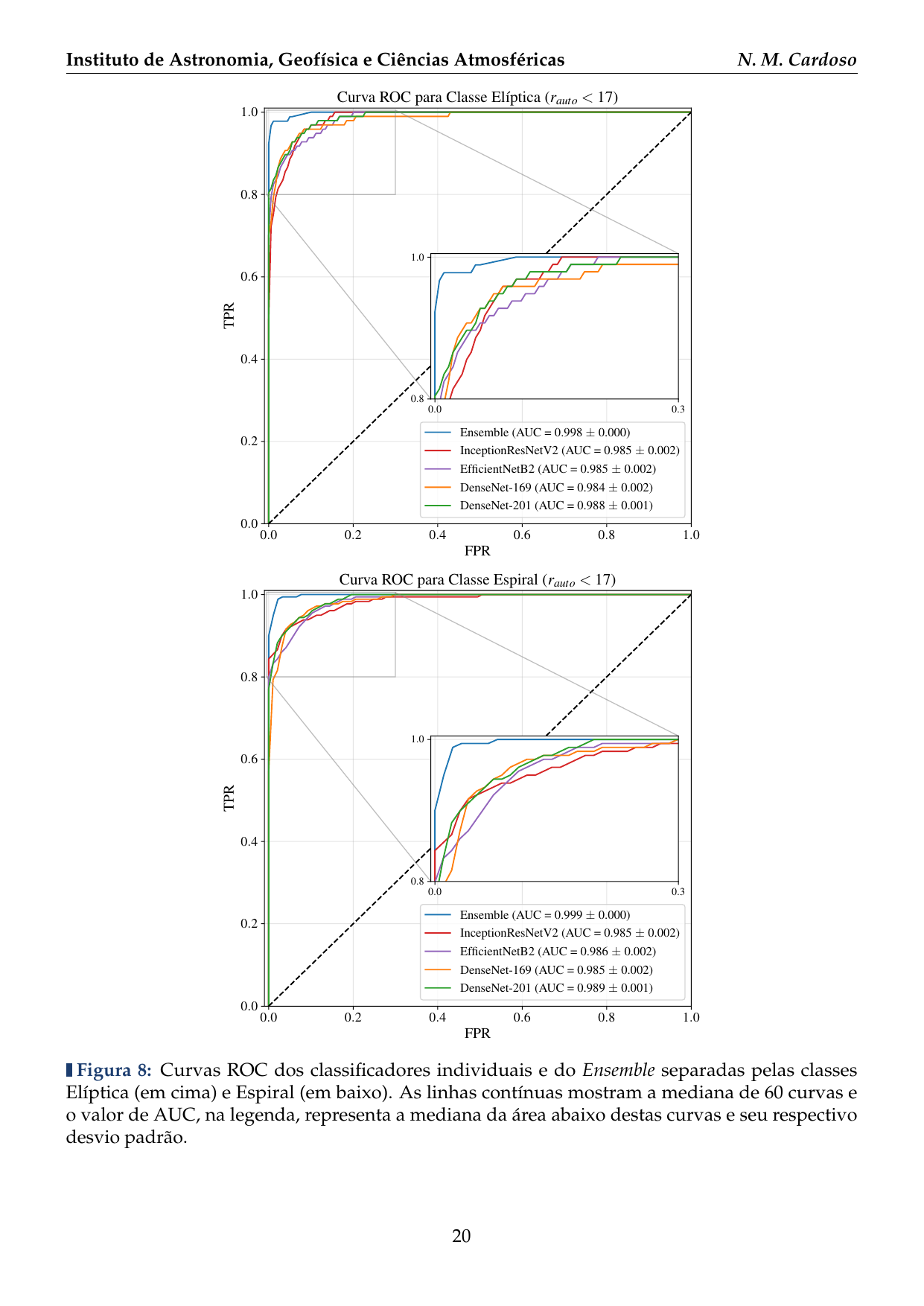
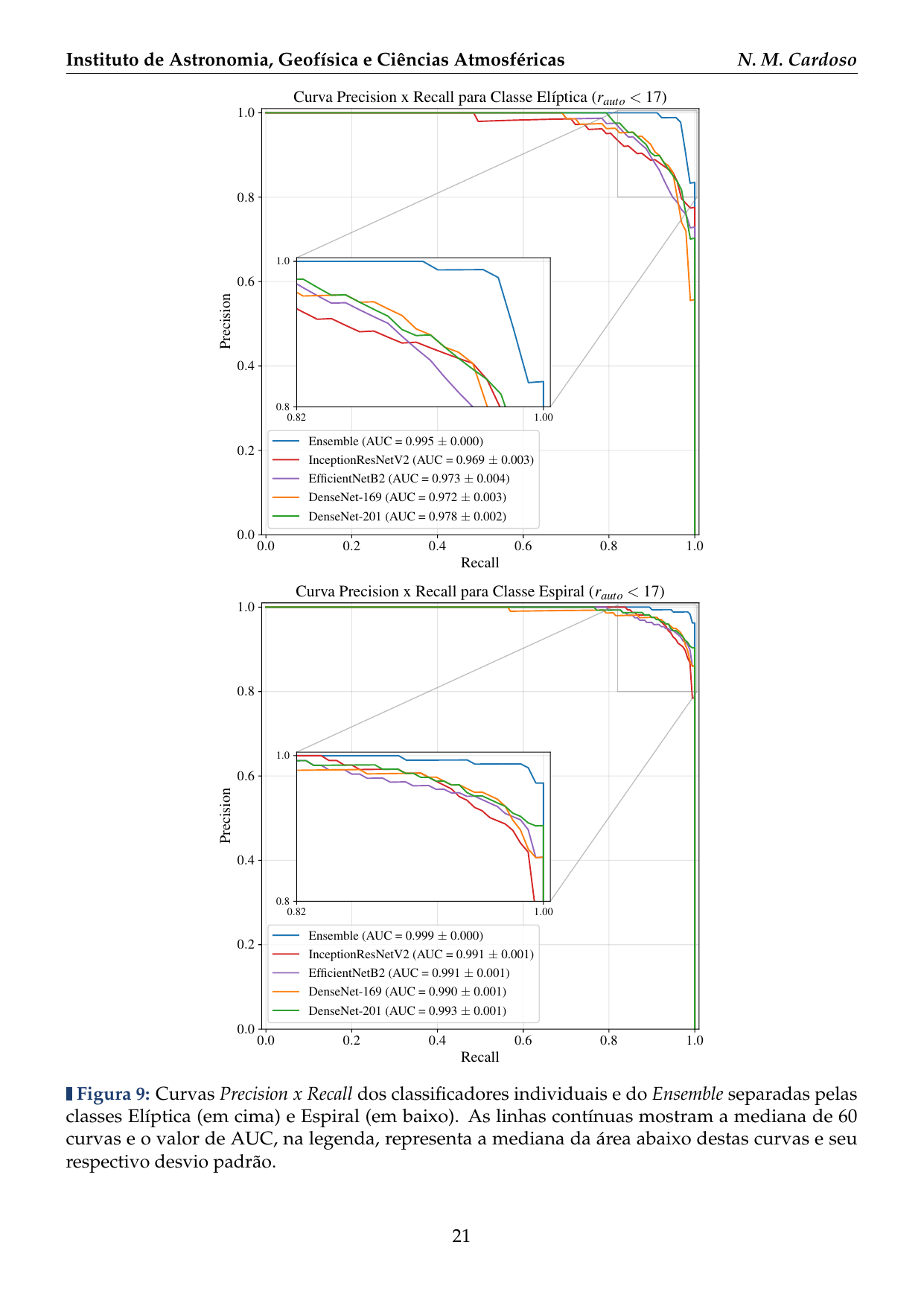
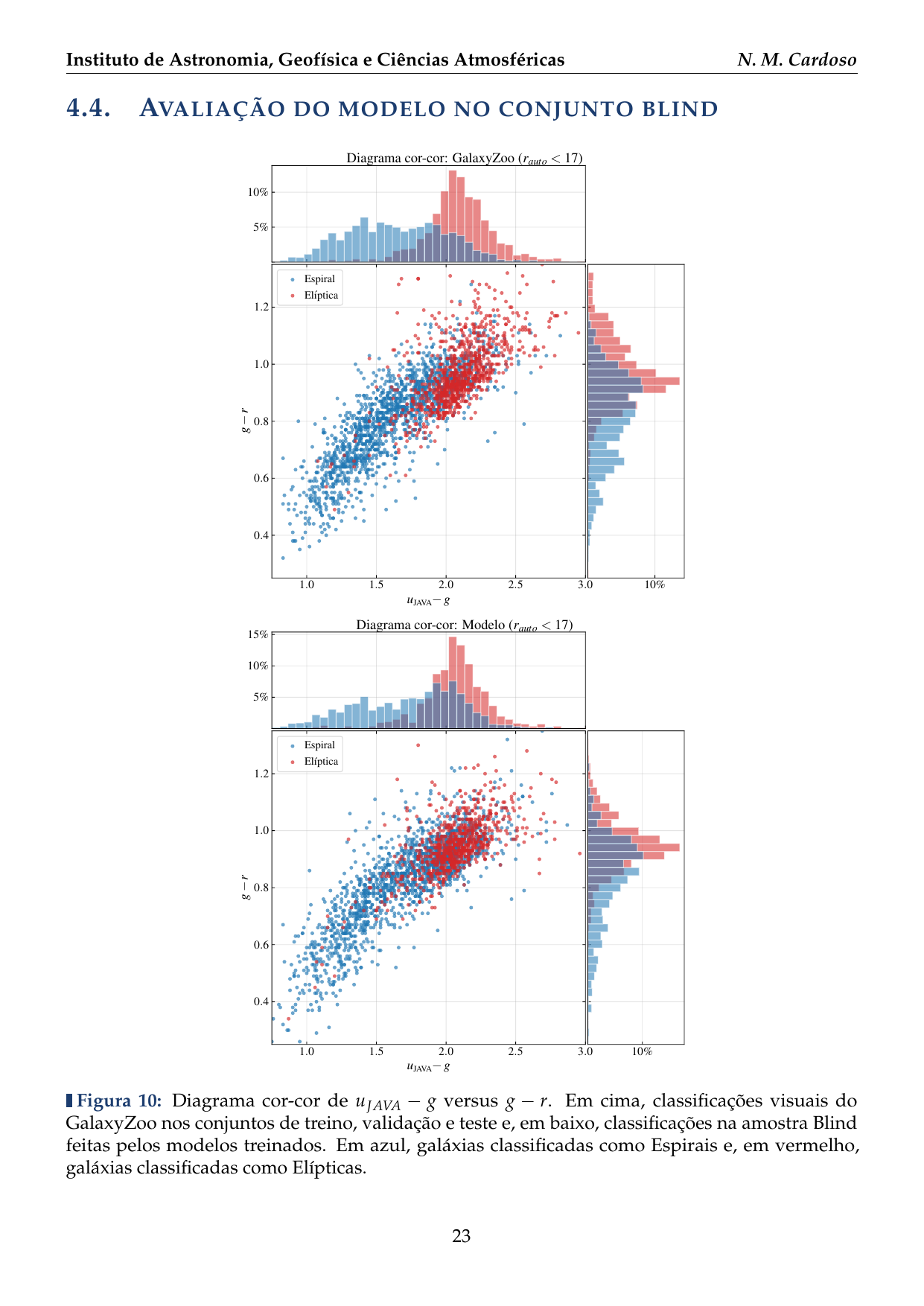
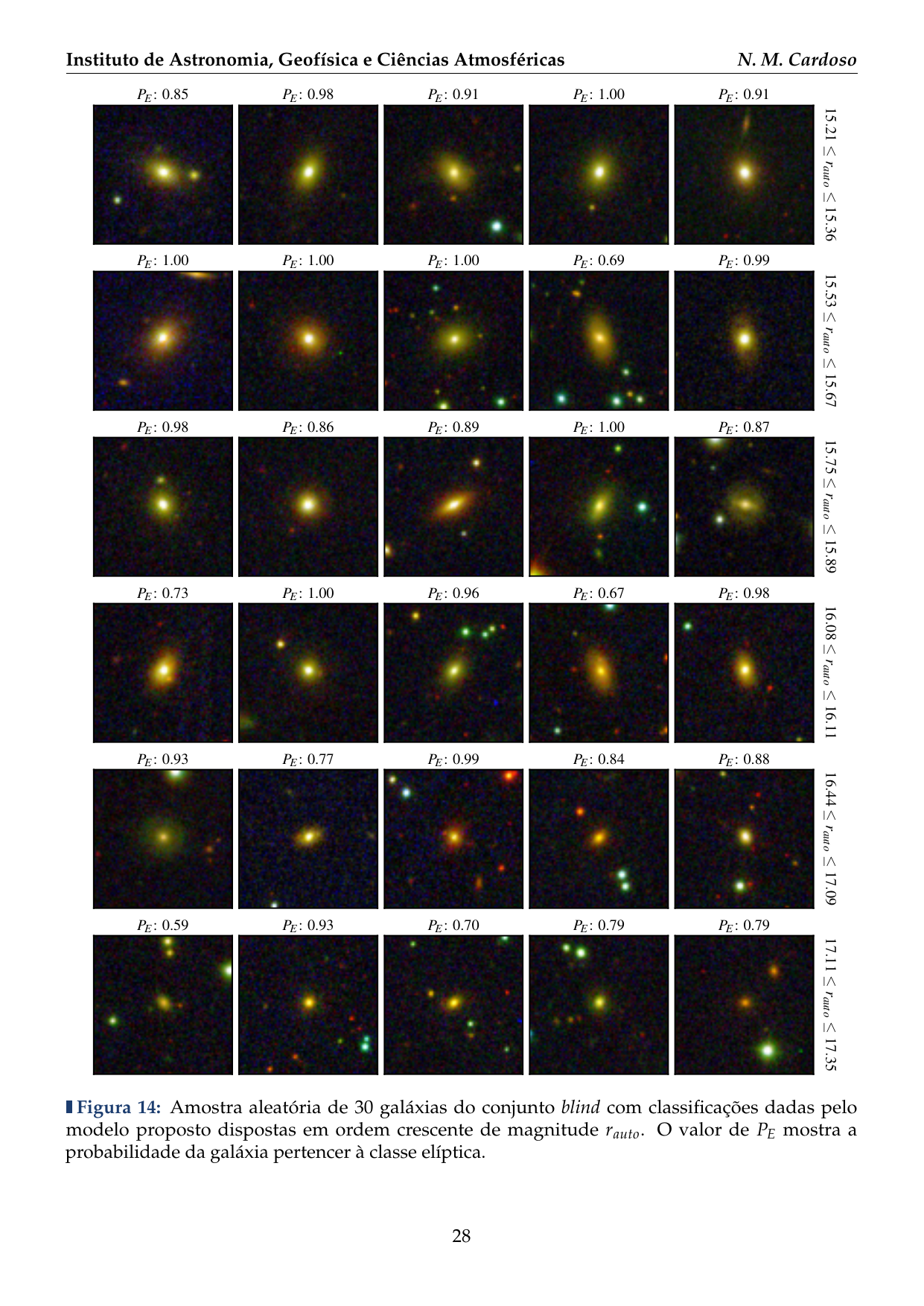
- 2016: Desenvolvimento de uma plataforma citizen science para coleta de dados em biodiversidade
- 2020: Classificação morfológica de galáxias do S-PLUS usando deep learning
- 2021: Classificação morfológica de galáxias usando conjunto de redes neurais convolucionais ⭐